自然界很多动物都会根据环境的变化来调节繁殖对策,产生不同数量的后代。动物种群的内在因素在调节繁殖对策方面也发挥着重要作用,比如种群密度、性比、亲缘关系和个体竞争强弱等都会影响所繁殖的后代数量。而后代的多少与种群的维持及动态密切相关,同时也是动物适应性的重要标志。人们对这种自然现象有所了解,但是对动物如何自主调控生殖策略的分子机制并不知晓。
飞蝗是研究密度依赖生殖策略的理想模型。根据种群密度不同,飞蝗存在群居型和散居型两种生态型。群、散飞蝗尽管基因型完全相同,但是它们采取的生殖策略有显著差异,即高密度的群居型飞蝗生殖力较低,以便将能量投入到长距离迁飞来寻找新的栖息地。而低密度散居型飞蝗采取较高的生殖力以产下更多的后代个体,来维系种群稳定性。两型飞蝗之间可以根据密度的变化相互转变,生殖对策也随之发生转换,进而导致产卵量发生变化。飞蝗这种适应种群密度变化而导致后代数量改变的机制是一个非常有挑战性的问题。
近日,康乐院士团队最新研究成果揭示了飞蝗适应种群密度变化而导致后代数量改变的分子机制。Piwi-interacting RNAs (piRNAs)是动物生殖系统中特异表达的一类非编码小RNA。群居型和散居型雌虫卵巢中piRNAs的表达差异非常明显。散居型飞蝗较高的产卵频率与piRNAs的高表达具有明显的关联性。特别是在散居型飞蝗卵小管尖端部位(原卵区和卵黄发生前期卵母细胞),piRNAs发生显著高表达。在散居型雌虫体内抑制飞蝗Piwi蛋白Liwi1、piRNAs或者piRNAs的靶标基因oo18 RNA-binding protein (Orb),都能够降低其产卵频率和卵块数。反之,在群居型雌虫体内过表达piRNAs,能够提高群居型雌虫的产卵频率和卵块数。有趣的是,piRNAs通过正调控模式影响靶标基因Orb的表达。目前国际上普遍认为Piwi/piRNAs通路经典的作用机制,是通过抑制转座子的转座来保证生殖系统内基因组的完整性。Piwi/piRNAs通路对蛋白编码基因的调控虽有报道,但大部分是抑制性的。那么散居型飞蝗中这些piRNAs是怎么实现对蛋白编码基因Orb转录本的正向调控呢?康乐院士团队研究发现,Liwi1/piRNAs作用的靶标是Orb内含子中一个不具有转座功能的残缺转座子“Copia”。飞蝗中Liwi1/piRNAs通过与剪切体蛋白U2AF35互作,促进内含子从前体mRNA中移除,导致产生更多的Orb成熟mRNA转录本,从而实现了Orb在散居飞蝗卵小管尖端的高表达。因此,piRNAs促进Orb前体mRNA剪切的这种非经典调控模式是导致散居型飞蝗高产卵频率和高产卵量的关键原因(图1)。
飞蝗生殖系统中为什么会发展出这样一种由Piwi/piRNAs通路介导的序列特异性的前体mRNA剪切模式呢?研究人员通过利用最新发展的针对微量细胞内RNA与蛋白互作研究的LACE-seq技术,发现在飞蝗卵小管尖端中,大量与卵母细胞分裂和发育密切相关的mRNA前体共同结合在Liwi1和U2AF35这两个蛋白上,提示生殖系统这种piRNAs介导的特殊剪切机制可能对生殖细胞的成熟至关重要。
Piwi/piRNAs参与的前体mRNA剪切模式是否在动物界普遍存在?研究人员进一步在果蝇及小鼠生殖细胞系中,验证了Piwi蛋白和剪切体蛋白的互作以及piRNAs介导的mRNA剪切机制的普遍性(图2)。这项研究也暗示了飞蝗中发现的Piwi/piRNAs介导的编码基因mRNA剪切新机制可能在其它动物调节繁殖对策中也是普遍存在的。
这项最新研究是继康乐院士团队发现piRNAs是群居型、散居型飞蝗中表达差异最大的一类小RNA (Wei et al, Genome Biology, 2009)之后,进一步证明了piRNAs对飞蝗生殖策略可塑性的调控机制,并揭示了Piwi/piRNAs介导的前体mRNA剪切对生殖功能相关基因的加工调控作用。该团队近期的另一项研究还发现,脑中的piRNAs通过调控神经肽NPF1的表达来控制飞蝗的取食量(Wang et al., EMBO Reports, 2022)。飞蝗piRNAs研究在生殖细胞和体细胞中同时取得的重要进展,既能为开发蝗灾控制方法提供新思路,又将飞蝗发展成为研究非编码RNA的重要模式昆虫。该成果以“piRNA-guided intron removal from pre-mRNAs regulates density-dependent reproductive strategy”为题于2022年4月26日在国际学术期刊Cell Reports杂志在线发表,中国科学院动物研究所助理研究员何静为该论文的第一作者,复旦大学基础医学院青年研究员魏园园和中国科学院动物研究所康乐研究员为共同通讯作者。
文章链接:http://doi.org/10.1016/j.celrep.2022.110593
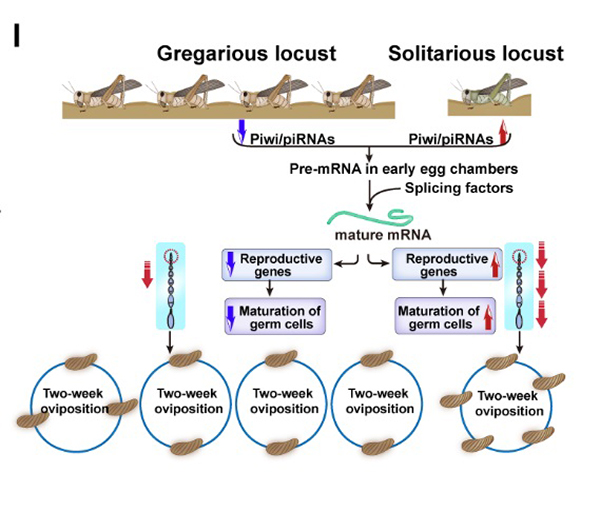 图1
图1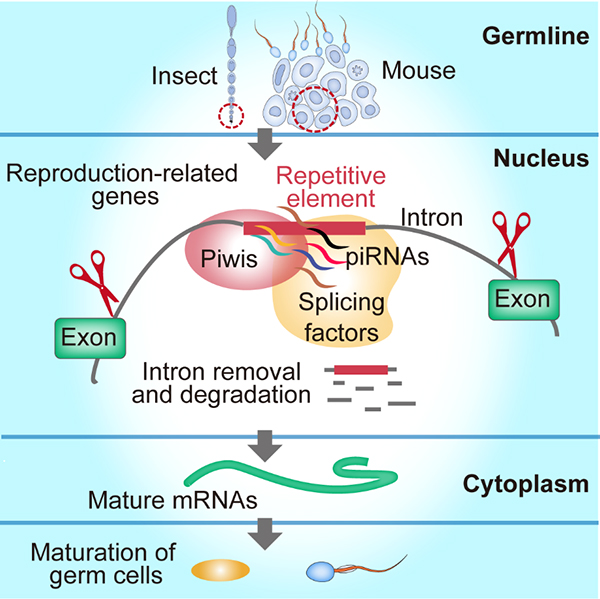 图2
图2