结直肠癌 (CRC) 是最常见的癌症之一,其发病率与特定的饮食模式有关。例如,西方饮食会增强肠道祖细胞的致瘤性并抑制抗肿瘤免疫。同样,高糖饮食和高血糖易患 CRC。此外,过量摄入动物蛋白,尤其是红肉,也会增加患CRC的风险。
生酮饮食(KD)是一种高脂肪、低碳水化合物,蛋白质和其他营养元素合适配比的饮食方案,起初是一种疗效确切的用于治疗儿童难治性癫痫的非药物疗法。
近日,来自美国宾夕法尼亚大学的科学家们在Nature发文报道称生酮饮食表现出强烈的肿瘤抑制作用。酮体 β-羟基丁酸 (BHB) 概括了生酮饮食的这些特性,能够减少了结肠隐窝细胞的增殖并有效地抑制了肠道肿瘤的生长。

研究者设计了具有确定的常量营养素来源、恒定蛋白质含量和不同脂肪与碳水化合物比率的小鼠饮食(图1a、b),并通过注射偶氮甲烷(AOM)和三个周期的葡聚糖硫酸钠(DSS)诱导结直肠癌(图1a),从而确定影响肠道肿瘤生长的饮食干预。
结果显示随着脂肪与碳水化合物的比例增加,肿瘤数量和大小受到抑制(图1c)。不仅如此,生酮饮食(KD)也能抑制Cdx2CreERTApcfl/fl CRC模型小鼠及已发生CRC小鼠的肿瘤进展,从KD饲喂恢复到正常饮食会导致CRC复发。这些结果表明,KD不仅能够预防小鼠CRC的发生,也能起到抑制CRC进展的效果。
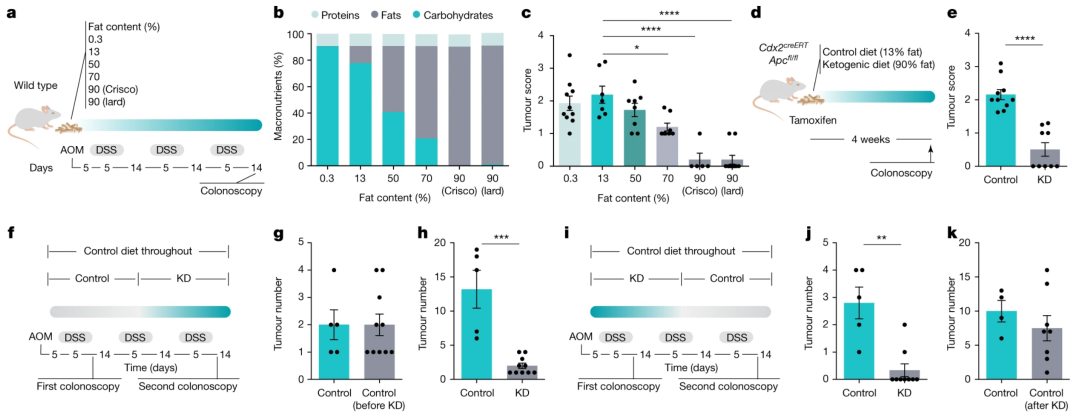
图1. a,AOM/DSS 处理小鼠的饮食暴露示意图。b , 膳食的常量营养素组成。c,喂食六种不同饮食的 AOM/DSS 治疗小鼠的肿瘤评分。d,Cdx2 CreERT Apc fl/fl小鼠的饮食暴露示意图。e ,喂食KD或对照饮食的Cdx2 CreERT Apc fl/fl小鼠的肿瘤评分。f-k,膳食暴露示意图 ( f , i ) 和基于结肠镜检查的肿瘤量化 ( g , h , j , k) 在治疗模型 ( f-h ) 和停止模型 ( i-k ) 中经 AOM/DSS 处理的 KD 喂养小鼠。
接下来,研究人员探索了 KD 对结肠肿瘤发展的影响的机制。
实验首先排除了热量摄入的差异。在 AOM/DSS 模型中,癌症进展是由免疫细胞驱动的,然而适应性免疫细胞在功能上并不是饮食保护作用所必需的。同时,NLRP3 炎性体也被证明和肿瘤发展机制无关。另外,他们使用了一个概括了肠上皮的干细胞衍生生长的类器官系统来研究 KD 对肠干细胞 (ISC) 的影响,表明饮食抑制了隐窝细胞的增殖。
生酮饮食(KD)能够刺激肝脏产生酮体乙酰乙酸 (AcAc) 和 酮体 β-羟基丁酸 (BHB),这是人体对饥饿的生理反应。为了确定KD对上皮生长的抑制作用是否由酮体介导,实验监测了在AcAc或BHB存在下培养的肠类器官(图2a)。尽管在 AcAc 存在下类器官的生长没有变化(图2b,c),但 BHB 以浓度依赖性方式减少了类器官的大小(图2d,e)。BHB 的这种作用也在肿瘤类器官中观察到(图2f,g)。除了诱导肝脏生酮外,KD通过降低全身葡萄糖水平和增强胰岛素敏感性赋予健康益处。
为了确定 BHB 补充和葡萄糖限制的相对贡献,我们使用不同的葡萄糖浓度用 BHB 处理类器官。结果表明葡萄糖限制和 BHB 补充可能通过不同的机制起作用。
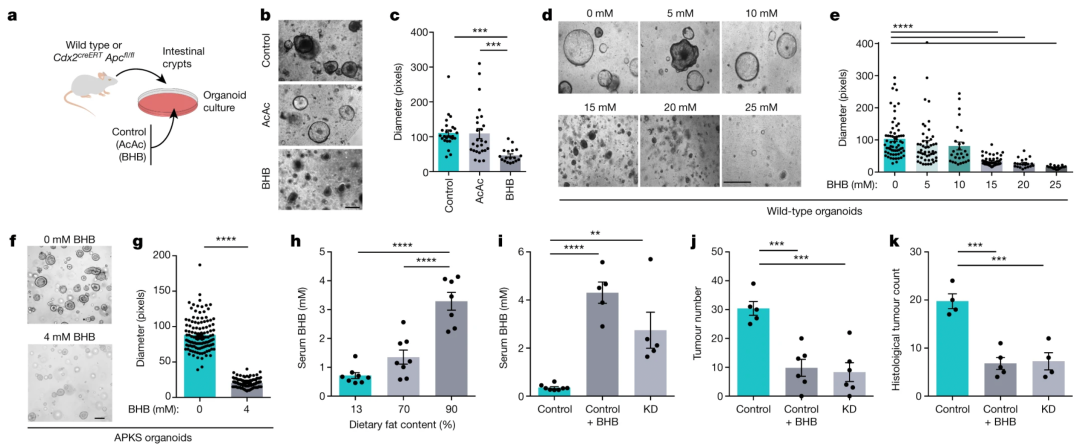
图2. a,用 AcAc 或 BHB 培养的类器官生长示意图。b、c 、暴露于 AcAc 或 BHB 的类器官的代表性图像 ( b ) 和量化 ( c )。d–g , 代表图像 ( d , f ) 和量化 ( e , g ) 野生型 ( d , e ) 和 AKS 类器官 ( f , g ) 与 BHB 一起培养。h,AOM/DSS 处理的小鼠中 BHB 的血清浓度,这些小鼠食用具有指定脂肪含量的饮食。i–k , 血清 BHB ( i)、基于结肠镜检查的肿瘤定量 ( j ) 和KD 喂养或 BHB 治疗的Cdx2 CreERT Apc fl/fl小鼠的组织学肿瘤计数 ( k )。
接下来,研究人员研究了酮体 β-羟基丁酸 (BHB) 介导的肿瘤抑制机制。他们对 BHB 处理过的类器官进行了 RNA 测序(RNA-seq),发现BHB 诱导了全局基因表达的显著变化(图3a)。其中一种抑制肿瘤发生的转录因子Hopx(表达集中于结肠隐窝基部,肠道干细胞分裂缓慢的标志物)的表达水平显著上升。
通过将来自 Hopx 缺陷小鼠和野生型同窝仔鼠的干细胞培养的类器官模型暴露于 BHB下发现,BHB 减少了野生型类器官的生长(图3c,d),而Hopx 缺陷类器官对 BHB 处理具有抗性(图3e,f)。因此,BHB 在野生型类器官中减少了上皮增殖,但在没有 Hopx 的情况下不受影响。Hopx 在类器官中的过表达足以减少野生型和肿瘤类器官的生长(图3g-j)。接受 KD 的小鼠在结肠组织中Hopx水平升高(图3k),在诱导结肠肿瘤后进一步加重(图3l)。这种效应是结肠独有的。而在 Hopx 缺陷小鼠和 KD 上的野生型同窝小鼠中诱导了 CRC后,虽然野生型组在 KD 喂养后肿瘤生长明显减少,但该饮食对缺乏 Hopx 的小鼠无效(图3m、n)。综上,KD 和 BHB 的肿瘤抑制作用需要Hopx的表达。
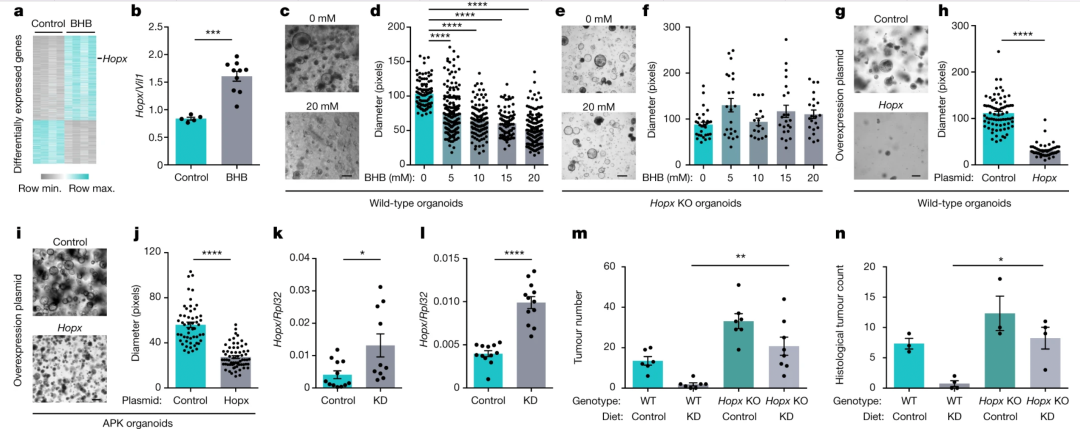
图3. a,BHB处理和对照类器官中差异表达基因的热图。b,BHB 处理的类器官中Hopx的表达。c-f ,用 BHB 处理的野生型(c,d)和 Hopx 缺陷型(e,f )类器官的代表性图像(c,e)和量化(d,f )。g–j,代表图像(g,i)和量化(h,j)野生型(g,h)和 APKS (i , j ) 过表达Hopx的类器官。k,l ,在稳态条件下 ( k ) 和肿瘤诱导后 ( l ) 喂食 KD 的小鼠中Hopx的结肠转录水平。m,n,基于结肠镜检查的肿瘤定量 ( m ) 和 KD 喂养的 Hopx 缺陷小鼠和对照的组织学肿瘤计数 ( n )。
文章同时讨论了BHB-HOPX 途径是否同样有效地抑制人肠上皮增殖(图4a)。事实上,BHB 减少了健康供体(图4b,c)和 CRC 患者中类器官的生长,表明肿瘤类器官对BHB 介导的抑制作用。与在小鼠中的发现相似,BHB 治疗导致人类类器官中Hopx表达升高(图4d)。
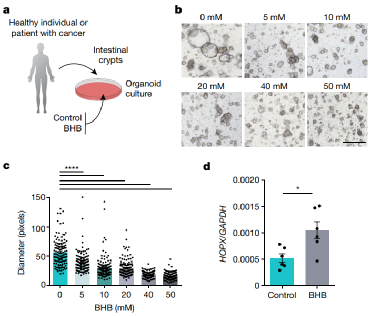
图4. a,从健康供体或用 BHB 或对照培养基培养的 CRC 患者获得的隐窝的肠道类器官生长示意图。b、c, 用指定浓度的 BHB 培养 10 天后,来自健康成年供体的类器官的代表性图像 ( b ) 和量化 ( c )。d,来自CRC患者的BHB治疗和对照类器官中HOPX的表达。
该研究结果提出了一个模型,其中肿瘤特异性基因表达模式决定了对特定饮食干预的反应程度。该模型提出了一种可能性,即通过调整膳食常量营养素组成以适应肿瘤类型以实现癌症预防和治疗的最佳结果,例如限制碳水化合物但含有足量膳食纤维的 KD 可以提供最佳的肠道肿瘤发生保护。尽管营养干预措施很有吸引力,但长期食用特定饮食很难维持,并可能导致全身副作用。饮食的肿瘤抑制作用可以通过补充代谢物来概括。BHB 介导的通路可能与手术、化学疗法或免疫疗法协同作用,并可能成为癌症治疗新支柱“代谢疗法”的一个例子。
参考文献:
[1] Oxana Dmitrieva-Posocco, Andrea C. Wong, Patrick Lundgren, et al. β-Hydroxybutyrate suppresses colorectal cancer. Nature 2022.
来自: 生物谷